Clique nas imagens para ampliar o seu tamanho.
Boa aventura no incrível mundo dos animais!

A classificação dos animais

1. Morfologia, Diversidade, biologia e Filogenia dos Porífera

Os Porifera (Latim: porus, poro; ferre, portador de;
daqui em diante poríferos) são metazoários multicelulares, geralmente com
simetria radial (ou assimétricos em alguns casos) e que possuem o corpo com
grande número de poros minúsculos, canais e câmaras que constituem um sistema
de filtragem/alimentação bem adaptado ao hábito de vida séssil. O corpo dos
poríferos possui forma de uma massa solida, a qual se encerra em um sistema de
canais e câmaras, no interior do qual entra água através de uma série de poros
finos, os óstios. A água passa por células especializadas, os porócitos, e é expelida
através de uma ou algumas aberturas maiores, os ósculos.
Alguns poríferos são eretos, outros se ramificam,
são lobados, pequenos ou, até mesmo, incrustantes. Não há nesses animais órgãos
nem tecidos distintos, entretanto, eles podem alcançar mais que dois metros de
diâmetro. Muitas espécies apresentam coloridos vivos devido à presença de
pigmentos em suas células dérmicas, as quais em alguns casos estão relacionadas
à toxicidade (aposematismo). As células dos
poríferos estão organizadas de maneira frouxa em duas camadas de células, uma
de cada lado (externo e interno), da matriz gelatinosa denominada meso-hilo. No
meso-hilo também são encontradas fibrilas e elementos esqueléticos. Externamente,
ele é revestido por um conjunto de células chamado pinacoderme, o qual dá
proteção aos poríferos, por exemplo, contra inimigos naturais. Internamente, ocorrem
as células denominadas arqueócitos, as quais são totipotentes, ou seja, que se
diferenciam em células que podem atuar em diferentes partes do corpo do animal.
Mas, geralmente essas células atuam na fagocitose de partículas e também na
digestão celular. No meso-hilo também estão inseridos os conócitos, células que
possuem colarinho e flagelo, os quais estimulam o fluxo de água e captam
oxigênio e partículas alimentares dissolvidas na água. Outras células que
ocorrem no meso-hilo são os miócitos, que são contrateis e ocorrem ao redor dos
ósculos ou poros (controle de abertura e fechamento); os esclerócitos que
secretam as espículas (caráter taxonômico); os espongiócitos que secretam fibras
de espongina do esqueleto; os colêncitos que secretam colágeno; e os lofócitos
que secretam colágeno, porém distinto morfologicamente daqueles dos colêncitos.
O meso-hilo também delimita uma cavidade interna, a espongiocele, onde a água
com oxigênio e partículas alimentares entra a partir dos óstios, é filtrada através
do movimento gerado pelo batimento dos flagelos dos conócitos e,
posteriormente, sai pelo ósculo.
Quanto ao modo de filtração, os poríferos podem ser
denominados asconoides (espongiocele flagelada), siconoides (canais flagelados)
e leuconoides (câmaras flageladas). Os
poríferos asconoides apresentam uma organização mais simples, sendo geralmente
pequenos e tubulares. Eles possuem coanócitos somente ao redor da parede
interna da espongiocele e um único ósculo. Os poríferos siconoides possuem canais
radiais forrados por coanócitos que desembocam na espongiocele. A água entra
pelos óstios dérmicos nos canais inalantes, passa para os canais radiais
através das prosópilas, é filtrada pelos coanócitos e, finalmente, vai para a
espongiocele, onde é eliminada pelo único ósculo. Já os poríferos leuconoides, os mais
complexos, possuem câmaras flageladas que recebem água dos canais inalantes, os
quais a impulsionam aos canais exalentes e que, eventualmente, a conduzem aos vários
ósculos. O último tipo de sistema (leuconoide) possibilita maior bombeamento da
água e, devido a maior superfície flagelada, também possibilita maior obtenção
de alimento e oxigênio. A maior parte das esponjas é do tipo leuconoide, sendo
a maioria dos poríferos calcários e todos os demais. Para ser ter uma dimensão,
o gênero Leuconia, por exemplo, é uma
leuconóide pequena (10 cm altura x 1cm diâmetro), para a qual estimou-se haver
81 mil canais inalantes e dois milhões de câmaras flageladas. Nos canais
inalantes a água passa por uma velocidade de 0,1 cm/s, já nas câmaras flageladas
a água desacelera a uma velocidade de 0,001 cm/s. Isso ocorre devido a soma dos
diâmetros das câmaras flageladas serem maiores que o diâmetro dos canais. Tal
adaptação possibilita uma ampla oportunidade para a captura de alimento pelas
células do colarinho. Depois disso, toda a água é exalada através de um único
ósculo a uma velocidade de 8,5 cm/s. Esponjas maiores podem filtrar 1,500
litros de água por dia.

Tipos de esponjas
A reprodução dos poríferos pode ser assexuada ou
assexuada. A reprodução assexuada geralmente ocorre por meio da formação de
brotos e pela regeneração seguida de fragmentação. Depois de atingir certo
tamanho, os brotos externos podem destacar-se da esponja parental e se mover a
partir da corrente de água até formar novas esponjas. Brotos internos ou
gêmulas são formados nas esponjas de água doce e em poucas de água marinha.
Nesses casos, os arqueócitos se juntam no meso-hilo e são envoltos por uma
camada de espongina incorporada com espículas silicosas. Quando a esponja
parental morre, as gêmulas sobrevivem e persistem dormentes durante períodos
inóspitos. Depois disso, as células das gêmulas saem pela micrópila, dando
origem a uma nova esponja. Outro modo de reprodução assexuada é a embriogênese
somática. Neste caso, quando o porífero é cortado em pequenos fragmentos, ou
seja, quando as células são inteiramente dissociadas e juntadas em pequenos
agregados, novos poríferos inteiros podem se desenvolver destes fragmentos.
Quanto à reprodução sexuada, os poríferos são
monóicos, sendo os oócitos e espermatozoides produzidos em momentos distintos.
Os espermatozoides surgem da transformação dos coanócitos e, em alguns casos,
os oócitos também se desenvolvem dos conócitos (e.g., Calcarea e Demospongiae).
A maioria dos poríferos é vivípara, mas há também especies ovíparas, as quais
os espermatozoides e oócitos são expelidos na água, onde fecundam e originam o
zigoto, que se desenvolve em larva livre-nadante. No caso dos vivíparos, os
espermatozoides são lançados na água e capturados pelo sistema de canal de
outro individuo. Os conócitos fagocitam o espermatozoide e, deste momento em
diante, os primeiros se tornam células carregadoras e protetoras. Elas levam o espermatozoide através dos
meso-hilo até os oócitos, onde ocorre a fecundação. Então, o zigoto recebe
nutrientes do porífero parental até que se desenvolva em uma larva ciliada
livre-nadante, que é liberada na água. Os embriões são livre-nadantes, mas os
adultos são sempre fixos, geralmente em rochas, conchas, corais ou outros
objetos submersos, às vezes também na areia. As larvas livre-nadantes da
maioria dos poríferos são parenquímulas de corpo mole.
Muitos animais como caranguejos, nudibrânquios e
peixes vivem como comensais ou parasitas dentro ou sobre os poríferos. Poríferos
maiores tendem a abrigar uma grande variedade de, principalmente,
invertebrados. Por outro lado, os poríferos crescem sobre muitos animais
viventes, como é o caso de moluscos, branquiópodes e corais. Há também
poríferos que são perfuradores de conchas ou rochas. Embora os poríferos tenham
poucos inimigos naturais, há alguns peixes que costumam raspar e se alimentar deles.
Quando injuriados, os poríferos podem se regenerar e restaurar as partes
perdidas. O processo de regeneração não implica em reorganização de todo o
animal, mas das partes feridas.
A maioria das mais de 10.000 espécies de poríferos
é distribuída em 22 ordens. As espécies marinhas representam mais que 95% do
filo. Elas são abundantes em todos os mares e profundidades e atualmente são classificados
em quatro classes, de acordo com seu esqueleto de espículas: Calcarea
(portadoras de espículas calcárias), Hexactinellida (espículas silicosas com
seis raios), Demospongiae (espículas silicosas e/ou espongina) e
Homoscleromorpha (sem espículas). Os
Calcarea possuem espículas retilíneas (monáxonas) ou possuem três ou quatro
raios. Elas ocorrem em águas rasas, tendem a ter 10 cm ou menos de altura e
podem ser asconoide, siconoide e leuconoide. Os Hexactinellida são de águas
profundas, variando de 7,5 cm a mais de 1,3 m em comprimento. Os Desmopongiae
contêm 95% das especies de poríferos viventes e incluem a maioria dos poríferos
maiores. As espículas podem estar ligadas umas as outras por espongina ou podem
estar completamente ausentes. Já os Homoscleromorpha, ocorrem tanto em águas
rasas quanto profundas com tamanhos variados, a maior parte deles leuconoide e
sem espículas.
Os poríferos
representam um grupo antigo, datado do Cambriano inferior ou Pré-cambriano.
Poríferos espongiformes ocuparam os recifes paleozóicos e, no período
Devoniano, ocorreu rápida diversificação das esponjas de vidro. No passado,
filogeneticistas indicaram a possibilidade dos poríferos terem derivado dos
coanoflagelados (protozoários que possuem colarinhos e flagelos) e, assim, os
coanoflagelados foram considerados grupo irmão dos metazoários. Contudo, devido
ao fato dos poríferos adquirirem seus colarinhos tardiamente em seu
desenvolvimento, esta hipótese foi muito questionada. Estudos recentes
envolvendo análises moleculares (RNA ribossômico) voltaram a sustentar a
hipótese dos coanoflagelados como grupo irmão e, ainda, reforçaram que os
poríferos são grupo irmão dos Eumetazoa. Por outro lado, outros estudos têm
sugerido que ao invés de Porifera, Ctenophora
seja o grupo irmão de todos os metazoários. Porifera seria grupo irmão
do clado Parahoxozoa Embora existam divergências sobre a classificação interna
dos poríferos, eles são considerados um grupo monofilético.
Fonte: maior parte – Hickman et al. Princípios integrados de zoologia.
11ª edição. Rio de Janeiro: Guanabara Koogan, 2004.
Poucos detalhes – Barnes et al. O invertebrados: uma síntese. São
Paulo: Ateneu, 2008.
2. Morfologia, Diversidade, biologia e Filogenia
dos Platelmintos e Nematódeos

Planária

Os Platyhelmynthes
(Grego: platy, chato; helminthes, vermes; daqui em diante platelmintos) são eumetazoários
com clivagem espiral, protostômios, multicelulares, triploblásticos, com simetria
bilateral e corpo achatado dorso-ventralmente, o qual possui as aberturas oral
e genital. Seu corpo é monomérico, embora possa ocorrer a formação de segmentos
a partir da zona de proliferação em algumas espécies. Nesses animais também há
polaridade defina em extremidade anterior, onde pode ocorrer cefalização, e
posterior. Espécies livre-nadantes geralmente possuem cabeça diferenciada e com
ocelos (fotorreceptor). O tamanho do corpo geralmente varia de um milímetro
(e.g. planárias) a mais que cinco metros em alguns casos (e.g. tênias).
A epiderme pode ser celular ou sincicial (núcleos
não separados por membranas celulares), mole e ciliada nos platelmintos de vida
livre ou revestida por cutículas e com ventosas para fixação no hospedeiro, nos
platelmintos parasitas. Não há cavidade corpórea (acelomados) e o esqueleto é
um tecido parenquimático, mas ocorre a formação de órgãos e tecidos. A parede
do corpo é composta por, principalmente, camadas de musculatura circular e
logintudinal. O sistema muscular é primariamente em forma de bainha e de origem
mesodérmica. A boca dos platelmintos é geralmente localizada na superfície
ventral, mas ela pode ocorrer na superfície médio-ventral em algumas espécies
de vida livre ou superfície terminal, quando presente em espécies parasitas. O
sistema digestório é incompleto do tipo gastrovascular, ou seja, tem boca,
faringe e intestino, mas não tem ânus. Ele também possui um fundo cego em forma
de saco ou, em alguns casos, é ramificado. Porém, ele pode ser ausente em
algumas espécies parasitas. Não há sistema circulatório e a osmorregulação e
excreção são feitas por dois canais laterais com ramos que apresentam
células-flama (protonefrídeos). A respiração ocorre principalmente por difusão.
O sistema nervoso consistindo em um par de gânglios anteriores com cordões
nervosos longitudinais conectados por nervos transversais e localizados no
mesênquima na maior parte das formas.

Há
aproximadamente 25 mil especies de platelmintos, sendo elas classificadas em:
Turbellaria (de vida livre), Trematoda (endoparasitas, principalmente de
vertebrados), Monogea (ectoparasitas, principalmente de peixes) e Cestoda
(endoparasitas). Os Tuberllaria mais bem conhecidos são as planárias, as quais
podem se regenerar e possuem os característicos rabditos e órgãos adesivos
duoglandulares, os quais possibilitam, respectivamente, a secreção de muco e
viscosidade que conferem proteção a esses animais. Os Turbellaria são, em sua
maioria, livre-nadantes e rastejantes. Sua movimentação se dá por meio de
deslizamentos ciliares ou ondas de contração muscular passando ao longo da
superfície ventral ou por todo o corpo.
São principalmente carnívoros, alimentando-se de pequenos crustáceos,
nematódeos, rotíferos e insetos, embora exista tambem saprófagos e parasitas.
Para tanto, possuem quimiorreceptores que os possibilita detectar o alimento a
pequenas distâncias. Também a faringe que é envolvida por uma bainha que se
abre posteriormente dentro da boca, através da qual ela pode estender-se. Uma
planária pode apreender sua presa com sua extremidade anterior, enrolar seu
corpo ao redor da presa, estender sua probóscide e sugar o alimento em pequenas
quantidades. A maior parte da digestão acontece de modo extracelular a partir
de enzimas proteolíticas liberadas no intestino. E, as demais partes do
alimento, são sugadas para o intestino, onde as células fagocitárias da
gastroderme completam a digestão (intracelular). Os Turbellaria são monóicos e,
durante a fase reprodutiva, desenvolvem órgãos masculinos e femininos, os quais
se abrem por um poro genital comum. Depois da cópula, um ou mais ovos
fertilizados e microscópios são formados e envolvidos em um casulo contendo
cada um várias células vitelinas dentro de uma casca. Os ovos se fixam em
rochas ou plantas por pequenas hastes e, então, os embriões emergem como formas
jovens que se assemelham aos adultos maduros. Em algumas formas marinhas, os
embriões desenvolvem-se em larvas ciliadas nadantes. Portanto, o
desenvolvimento é direto.
Devido ao hábito parasitário, os Trematoda possuem
varias glândulas de penetração ou glândulas para produção de material para
cistos. Alguns, como a Fasciola hepática
e Schistosoma mansoni, são
importantes parasitas dos humanos. Nos Trematoda ocorre um dos ciclos mais
intrigantes da natureza, em que a reprodução sexuada ocorre dentro do
hospedeiro definitivo, os ovos saem através das fezes do hospedeiro e eclodem
em larvas miracídias na água. Depois disso, essas larvas precisam encontrar o
hospedeiro intermediário (caramujo), onde se transformam em esporocistos,
reproduzem-se assexuadamente e dão origem as larvas cercárias, as quais
penetram a pele humana (hospedeiro definitivo). Já os Monogea geralmente usam
um único hospedeiro e possuem as larvas oncomiracídeas, as quais possuem grandes
ganchos na extremidade posterior (opistáptor). Tanto os Monogea quanto os
Trematoda pastam nas células de seus hospedeiros, alimentando-se de seus restos
e fuidos corpóreos. Como visto, nelas podem ocorrer numerosos estágios larvais
secundários e, portanto, seu desenvolvimento é indireto. Finalmente, os Cestoda
se diferenciam dos demais nematódeos por possuírem corpos longos e mais
achatados nos quais existe uma série de conjuntos de órgãos reprodutivos (proglótide)
e o escólex, um órgão de fixação. Eles não possuem sistema digestório, vivendo
totalmente à custa de seus hospedeiros, incluindo os humanos (e.g. Taenia spp.). A reprodução dos Cestoda
se assemelha muito as dos demais parasitas, Monogea e Trematoda.

Estudos filogenéticos envolvendo a análise de DNA
ribossômico, padrões embrionários de clivagem, origens do mesoderma e
estruturas do sistema nervoso, têm sugerido que os Acoela, um dos táxons dentro
de Platyhelminthes, não fazem parte do filo. Sendo assim, Platyhelminthes seria
polifilético. As relações entre os Platyhelminthes com Nemertea e
Gnathostomulida não estão claras, mas aparentemente os Platyhelminthes pertecem
ao clado Spirallia (clivagem espiral), dentro Protostomia. Dentro de Platyhelminthes,
parece claro que Turbellaria, mesmo excluindo Acoela, é parafilético.
Já os Nematoda (Grego: nema, filamento; eidos,
forma; daqui em diante nematoides) são metazoários multicelulares, protostômios,
triploblásticos e o seu padrão de clivagem embrionária é determinado, com um
arranjo de células formando um “T” no estágio de quatro blastômeros. A simetria
é bilateral, mas com uma tendência a uma simetria radial ao redor do eixo
longitudinal. A maioria dos nematódeos possui menos de cinco centímetros, sendo
muitos microscópicos, embora alguns poucos possam ter mais de um metro de
comprimento. O corpo é cilíndrico e delgado, revestido por uma cutícula dura e
resistente. A parede do corpo possui apenas fibras musculares longitudinais. A
epiderme possui os núcleos das células restritos a quatro cordões dispostos
mediano-dorsalmente, ventralmente e lateralmente. O espaço interno do corpo
representa um pseudoceloma não revestido, geralmente derivado da blastocele. Os
órgãos internos geralmente possuem número definido de células, o que é
denominado constância celular. Grande parte da cavidade corpórea dos nematódeos
é preenchida com um par de órgãos reprodutivos, ovários e testículos, os quais
são dispostos serialmente, às vezes enrolados. A cavidade interna do corpo
contém fluido corpóreo sempre em alta pressão, o que possibilita mudanças na
forma do corpo devido ao fato da musculatura longitudinal trabalhar contra a
pressão (esqueleto hidrostático). Neste sistema de alta pressão, a secção
transversal do corpo é sempre circular. O sistema digestório é completo, sendo
um tubo reto com musculatura partindo da boca ao ânus, de uma extremidade a
outra do corpo. Não há órgãos circulatórios ou respiratórios, e a excreção
ocorre a partir de uma ou mais células glandulares maiores (sem protonefrídeos).
A respiração ocorre por difusão. O sistema nervoso possui nervos longitudinais,
sendo um anel nervoso ao redor do esôfago ligado a seis nervos anteriores e
outros seis posteriores.

Há aproximadamente 15 mil especies de nematódeos, mas
as estimativas sugerem, pelo menos, um milhão. Eles são divididos em:
Adenophorea e Secernentea, sendo a maior parte deles parasitas, inclusive de
humanos (e.g., Ascaris lumbricoides e
Ancylostoma duodenale). Mas, há
especies de vida livre, as quais são predadoras de outros nematódeos,
bacteriófagas, fungívoras, suspensívoras, detritívoras ou herbívoras. Os Adenophorea
não possuem fasmídeos, ou seja, órgãos sensoriais caudais, porem ele têm anfídeos
de formas variadas (raramente em forma de poro), que são os órgãos sensoriais
anteriores. Eles geralmente são de vida livre. Já os Secernentea possuem fasmídeos
e anfídeos em forma de poro e a maior parte deles é parasita.
Nematódeos são encontrados no mar, na água doce e
no solo. Eles são dioicos, com o macho sendo menor que a fêmea. As gônadas são
contínuas, com ductos reprodutivos em ambos os sexos. Diferente dos platelmintos,
nos nematódeos não ocorre reprodução assexuada ou regeneração. A fecundação é
interna, dando origem a ovos microscópicos, os quais são revestidos por casca
quitinosa. O desenvolvimento do embrião é determinado e indireto. A larva passa
por várias mudas, representando, em alguns casos, o estágio infectante.
Os nematódeos foram agrupados com Rotirefa,
Gatrotricha, Kinorhyncha e Nematomorpha em um único filo chamado Aschelminthes,
devido a caracteres morfológicos. Recentes estudos envolvendo análises
moleculares de RNA ribossômico e desenvolvimento embrionário sugerem que esses
táxons podem estar agrupados de maneira diferente, em Lophotrochozoa e
Ecdysozoa. Nesse caso, os nematódeos seriam mais aparentados aos Nematomorpha,
dentro dos Ecdysozoa, no clado Nematoida (apenas com musculatura longitudinal,
espermatozóides sem flagelo e cloaca em ambos os sexos). Embora tudo isso,
estudos têm sugerido que Nematoda deve ser monofilético.
Fonte: maior parte – Barnes et al. O invertebrados: uma síntese. São
Paulo: Ateneu, 2008
Poucos detalhes – Hickman et al. Principios integrados de zoologia.
11ª edição. Rio de Janeiro: Guanabara Koogan, 2004.
Storer et al. Zoologia Geral. 6ª edição. São Paulo: Companhia Editora,
2003.
3. Morfologia, Diversidade, biologia e
Filogenia dos Anelídeos

Poliqueto


Minhoca
Os Annelida (Latim: annelus, diminutivo de anulus,
um anel; daqui em diante anelídeos) são eumetazoários protostômios, com
clivagem espiral, desenvolvimento em mosaico, multicelulares, triploblásticos, com
simetria bilateral, cefalização e corpo alongado. Na maior parte deles o corpo
é conspicuamente segmentado interna e externamente (metamerizado). Os segmentos
(ou metâmeros) são em geral semelhantemente arranjados em uma série linear e
delimitados externamente por sulcos circulares (ânulos). A parte anterior do
corpo apresenta um prostômio e a posterior um pigídio, ambos não sendo
considerados segmentos. Porém, é comum a fusão dos segmentos anteriores com o
prostômio para formar a cabeça. A segmentação dos anelídeos é estabelecida
durante o desenvolvimento, a partir de zonas mesodérmicas de crescimento
pareadas e de um anel ectodérmico correspondente à frente do pigídio, região
posterior, através da qual a endoderme se abre para o exterior. Novos segmentos
são formados na face anterior do pigídio, de modo que o último segmento a ser
formado é sempre o mais posterior. Esses animais podem continuar a adicionar
segmentos ao longo de toda sua vida, embora um número definitivo seja
geralmente alcançado na maturidade.
O corpo dos anelídeos é revestido por uma cutícula
fina e úmida sobre um epitélio colunar que contém glândulas unicelulares e
células sensitivas. A parede do corpo e o sistema digestório possuem camadas de
músculos circulares e longitudinais. Seus apêndices são cerdas quitinosas
filiformes diminutas, variando em quantidade por segmento dependendo do táxon.
Por exemplo, Oligochaeta possui um apêndice por segmento, enquanto Polychaeta
geralmente possui dois e Hirudinea não possui. O celoma (esquizocele) é bem
desenvolvido na maior parte dos anelídeos, com exceção aos Hirudinea. Ele é
forrado pelo peritônio, o qual reveste todos os órgãos internos e forma os
mesentérios dorsal e ventral. Os
peritônios dos segmentos adjacentes unem-se para formar os septos, que dividem
o corpo internamente e geralmente são comuns nos Oligochaeta e Polychaeta. O sistema
circulatório é fechado, com vasos sanguíneos longitudinais, ou seios
celomáticos, com ramos laterais em cada segmento. O plasma sanguíneo muitas
vezes possui amebócitos livres e pigmentos respiratórios dissolvidos (e.g.
hemoglobina, hemeritina ou clorocruorina). Por serem animais de corpo mole com
um grande celoma cheio de líquidos, esses animais possuem um esqueleto
hidrostático em que a contração dos músculos da parede do corpo atua contra o
líquido interno, possibilitando o movimento. Assim, a contração da musculatura
longitudinal da parede do corpo determina o encurtamento e o consequente
aumento no diâmetro do corpo, ao passo que a contração dos músculos circulares
causa alongamento e afilamento do corpo desses animais.
O sistema digestório dos anelídeos é completo,
tubular e estende-se por todo o comprimento do corpo. A respiração desses
animais ocorre pela epiderme, parapódios ou por brânquias, a última geralmente em
espécies tubícolas. O sistema excretor possui um par de nefrídeos por segmento,
cada um deles removendo as excretas do celoma e corrente sanguínea diretamente
para o exterior. Já o sistema nervoso tem um cérebro formado por um par de
gânglios cerebrais com conectivos, os quais são ligados a um cordão nervoso
duplo posicionado ventralmente e que se estende por todo o comprimento do
corpo. Há também um gânglio e pares de nervos laterais em cada segmento. Esses
animais percebem o ambiente a partir de células e órgãos sensitivos para o
tato, paladar (papilas gustativas) e fotorrecepção, tendo olhos com lentes em
algumas espécies.

Corpo de uma minhoca
Há cerca de 10 mil espécies de anelídeos descritas,
mas esse número pode chegar a e 75 mil dependendo da literatura consultada,
sendo elas divididas em três principais grupos: Polychaeta, Oligochaeta e
Hirudinea. O tamanho da maior parte dos anelídeos varia de cinco a dez
centímetros de comprimento, embora existam aqueles menores do que um milímetro
e outros maiores que quatro metros (e.g. minhocoçus gigantes da região
tropical). Alguns possuem coloridos vivos em tons de verde e vermelho, enquanto
que outros são iridescentes ou opacos. Desempenham importante papel ecológico,
geralmente servindo como alimento para peixes, crustáceos, hidrozoários, entre
outros (Polychaeta). Mas, por outro lado, podem ser importantes parasitas
(Hirudinea) ou predadores (Hirudinea, Polychaeta), algumas com importância
medicinal (Hirudinea: Hirudo medicinalis),
e outras possibilitando a oxigenação do solo (Oligochaeta).
Os Polychaeta se diferenciam dos demais anelídeos
por possuírem uma região cefálica bem diferenciada com órgãos sensoriais
especializados, e também pela maioria dos segmentos possuírem apêndices pares
denominados parapódios, os quais portam feixes de cerdas. Eles provavelmente
representam dois terços do número total de espécies de anelídeos, sendo eurialinos
e, portanto, sua maior parte marinha. Boa parte das espécies é bentônica,
embora existam algumas pelágicas que fazem parte da comunidade planctônica. Vivem
sob pedras, em fendas de corais, conchas abandonadas ou escavam o lodo e areia.
Algumas constroem seus próprios tubos sobre objetos submersos, mas há aquelas
que utilizam os tubos ou galerias de outros animais. Muitas espécies passam a maior
parte de suas vidas em tubos ou galerias, especialmente aquelas com elaborados
dispositivos para alimentação e respiração. Por exemplo, os representantes do
gênero Amphitrite possuem tentáculos
e brânquias voltadas para fora de seus tubos, possibilitando que elas capturem
oxigênio e fragmento de matéria orgânica sem precisar sair. Outras especies
nadam ativamente, cavam, rastejam ou saem de seus tubos para capturar alimento
e para se reproduzir. Os Polychaeta não possuem órgãos reprodutores permanentes
e geralmente são dióicos. As gônadas são projeções temporárias do peritônio que
liberam gametas para o celoma. Os gametas vão para o exterior através de
gonodutos, metanefrídios ou pela ruptura da parede do corpo. A fecundação é
externa com a posterior formação de um estágio larval trocófora. A reprodução
assexuada por brotamento pode ocorrer tanto em alguns Polychaeta, quanto em
alguns Oligochaeta.
Os Oligochaeta e Hirudinea são geralmente
associados ao solo e águas continentais. Eles se diferenciam dos Polychaeta
principalmente pela presença do clitelo, um órgão externo que secreta muco. Algumas
espécies de Oligochaeta e Hirudinea de água doce enterram-se no lodo ou areia,
enquanto outras o fazem entre a vegetação submersa. Boa parte dos Oligochaeta é
terrestre ou dulciaquícola e evitam a luz, incluindo as familiares minhocas. As
suas cerdas podem ser longas, curtas, restas, curvas, rombas, pontiagudas,
dispostas isoladamente ou em feixes, sempre em menor número que os Polychaeta. A
maioria dos Oligochaeta é saprófaga. Eles são monóicos, com dois pares de
testículos e um par de ovários. Os espermatozoides amadurecem nas vesículas
seminais, passam para os funis, para os dutos e depois chegam aos poros
genitais no segmento 15, onde são expelidos durante a cópula. Já os óvulos são
liberados na cavidade celomática, onde os funis ciliados dos ovidutos os
transportam para o meio externo através dos poros genitais femininos no
segmento 14. No segmentos nove e dez existem dois pares de receptáculos
seminais que recebem e armazenam o esperma traçado entre os parceiros durante a
cópula. Quando pareiam, aproximam as partes ventrais e se mantém unidos pelo
muco secretado pelo clitelo e por cerdas especiais que penetram o corpo do
parceiro. Depois da cópula, cada parceiro secreta um tubo mucoso formando um
casulo que ao se deslizar para frente, recebe os óvulos provenientes do oviduto.
A fecundação ocorre no casulo e, quando este se desprende, suas extremidades se
fecham. A embriogênese se da no casulo com posterior formação de um jovem que
eclode com forma semelhante ao adulto. Portanto, o desenvolvimento é direto e
sem metamorfose. Já os Hirudinea possuem muitas espécies predadoras, enquanto que
outras se especializaram em perfurar e sugar o sangue de tecidos moles de seus
hospedeiros. Para o hábito parasitário dos Hirudinea desenvolveu neles um par
de ventosas, as quais possibilitam a fixação ao hospedeiro/presa. Assim como os
Oligochaeta, os Hirudinea são monóicos com fecundação cruzada. Porém, o esperma
é transferido pelo pênis ou por transferência de espermatóforo, o qual penetra
a epiderme (impregnação hipodérmica). Como nos Oligochaeta, um casulo onde a
embriogênese ocorre é formado, mas os casulos são enterrados e presos a objetos
submersos ou no solo úmido.
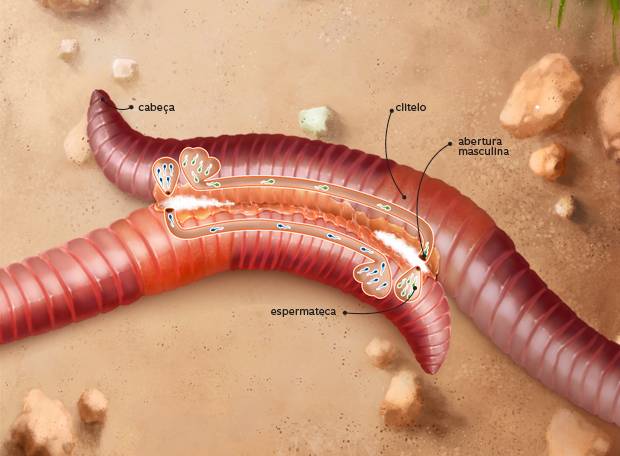
Esquema da reprodução em minhocas

Reprodução em minhocas
Os anelídeos estão entre os três maiores filos de
protostômios, juntamente com Mollusca e Arthropoda. Estudos filogenéticos apontam
que há semelhanças entre o desenvolvimento embrionário destes três filos,
sugerindo estreita relação entre eles. Algumas literaturas têm sugerido que os
três filos formam um grupo irmão dos Platyhelminthes. Ainda, a metameria
representa uma condição homóloga entre os anelídeos e os Arthropoda, os
colocando em um taxon chamado Articulata. Contudo, análises moleculares indicam
que os Mollusca e os anelídeos estão no clado chamado Lophotrochozoa, do qual
os Arthropoda não fazem parte. De maneira geral, os anelídeos são vistos como
um grupo monofilético, dentre dos quais, os Polychaeta são, por conveniência,
divididos em sedentários e errantes, e os Hirudinea e Oligochaeta formam um
clado chamado Clitellata.
Fonte: maior parte – Hickman et al. Principios integrados de zoologia.
11ª edição. Rio de Janeiro: Guanabara Koogan, 2004.
Poucos detalhes –Barnes et al. O invertebrados: uma síntese. São
Paulo: Ateneu, 2008
Storer et al. Zoologia Geral. 6ª edição. São Paulo: Companhia Editora,
2003.
4. Morfologia, Diversidade, biologia e
Filogenia dos Insetos

Formiga
Insetos é o nome popular dado a todos os Hexapoda,
ou seja, animais segmentados com seis pernas. Eles são eumetazoários
multicelulares, triploblásticos, protostômios, com clivagem geralmente
superficial e simetria bilateral. O corpo dos insetos constitui-se de uma
sequência de metâmeros que se fundem ao longo do desenvolvimento e formam
regiões denominadas tagmas (cabeça, tórax e abdômen). Os metâmeros que se
fundem e revestem a parte dorsal do corpo representam o tergo, os que revestem
a parte lateral representam a pleura e os que revestem a parte ventral formam o
esterno. Formados os tagmas, em cada um deles ocorrem funções diferenciadas. A
cabeça é principalmente relacionada à parte sensorial e de alimentação, o tórax
é relacionado à inserção da musculatura responsável pela locomoção e/ou voo, e
o abdômen é relacionado principalmente às funções de digestão e reprodução.


Morfologia externa de um besouro
Há a hipótese de que a cabeça dos insetos seja
resultado da fusão dos seis ou sete primeiros metâmeros. Nela ocorre o aparato
bucal constituído por mandíbulas, maxilas, lábio (segundas maxilias fundidas) e
hipofaringe, adaptado para mastigar (e.g. Coleoptera), sugar (e.g. Hemiptera) e
lamber (e.g. Diptera). Na cabeça também podem estar presentes dois olhos
compostos, três ocelos e um par de antenas (exceto Protura). O tórax é dividido
em três metâmeros, ocorrendo em cada um deles a inserção de um par de pernas,
as quais são constituídas de coxa, trocânter, fêmur, tíbia e tarso. Nos
Pterygota, é nos dois últimos segmentos do tórax que se inserem os dois pares
de asas, embora existam ordens com um par (e.g. Diptera) ou nenhum (e.g. Siphonaptera;
Apterygota). No plano básico dos insetos, o abdômen geralmente possui onze
metâmeros, embora possa haver menos. Neste tagma podem ocorrer os cercos
(sensoriais) e a terminália (onde estão dispostas as genitálias).
O sistema digestório dos insetos é completo, com
estomodeu (boca, faringe, glândulas salivares, esôfago, papo e moela),
mesênteron (estômago e cecos gástricos) e proctodeu (intestino, reto e ânus).
Parte da digestão ocorre no papo, enquanto o alimento se mistura com enzimas da
saliva. A absorção do alimento ocorre principalmente no mesênteron e os cecos aumentam
a área de absorção e digestão. No proctodeu ocorre pouca absorção de alimentos,
mas grande parte da reabsorção da água. A excreção ocorre a partir dos túbulos
de Malpighi (exceto Collembola), os quais são túbulos finos de fundo cego
ligados ao mesênteron e proctodeu, em número variável. Os túbulos de Malpighi
possibilitam a excreção de ácido úrico (insolúvel em água) a partir de reações
químicas envolvendo a secreção ativa de potássio. Assim, as fezes e urina dos
insetos são praticamente secas, o que os confere boa vantagem em ambientes
áridos.

As trocas gasosas da maior parte dos insetos
ocorrem por traqueias ramificadas (sistema traqueal), as quais constituem um
conjunto de tubos internos. Brânquias ocorrem em insetos aquáticos,
especialmente no estagio imaturo (e.g. Odonata). O ar chega e sai desses tubos
a partir de orifícios externos situados no tórax e no abdômen, denominados espiráculos.
O sangue dos insetos, hemolinfa, não tem função respiratória. Ela ocupa todos
os espaços da cavidade geral do corpo (hemocele). Assim, o sistema circulatório
dos insetos é aberto e o único órgão presente é o vaso dorsal. O vaso dorsal
estende-se da região posterior do abdômen até a cabeça, onde passa por baixo do
cérebro e abre-se acima do esôfago. Ele é diferenciado em uma aorta anterior,
um tubo contínuo e uma porção posterior expandida (coração, parte pulsátil). A
hemocele é dividida em três cavidades: o seio pericárdico (dorsal), o seio
perineural (ventral) e o seio perivisceral (central). Nessas cavidades e próximo
ao coração ocorrem os ostíolos, os quais possibilitam a entrada da hemolinfa
novamente no vaso dorsal depois de liberadas na hemocele. Desse modo, o coração
geralmente impulsiona a hemolinfa da região terminal do abdômen em direção à
cabeça. Depois de passar pelo corpo a hemolinfa volta ao coração a partir dos
ostíolos.
Já o sistema nervoso dos insetos é constituído por
um cérebro dividido em três regiões: o protocérebro (especialmente visão),
deutocérebro (especialmente percepção antenal) e tritocérebro (especialmente
alimentação). Além disso, há gânglios supra e subesofágicos ligados ao cordão
nervoso ventral duplo, com um par de gânglios por metâmero. Além dos olhos e
ocelos, o sistema sensorial dos insetos conta com quimiorreceptores para olfato
nas antenas, para paladar perto da boca e pêlos táteis variados. Alguns insetos
possuem espinhos ou outras estruturas que produzem e percebem sons (e.g.
Orthoptera). Outros têm bioluminescência (e.g. luciferina em Lampyridae). Não
ocorrem estatocistos nos insetos.
São descritas aproximadamente um milhão de espécies
de insetos, embora algumas estimativas cheguem a sugerir que ocorram até 30
milhões. Assim, eles são os organismos mais diversos e também abundantes na
Terra (60% de todas as especies conhecidas e 2/3 da biomassa terrestre). São organismos que variam muito em tamanho, de
menos que um milímetro (mm) a mais que 35 centímetros (cm), embora a maior
parte tenha de dois mm a quatro cm. As
cores variam de crípticas (e.g. Lepidoptera: mariposas), apostemáticas (e.g.
Lepidoptera: borboletas) a iridescentes (e.g. Hymenoptera e Coleoptera). O
corpo pode ser alongado (e.g. Phasmatodea), arredondado (e.g. Coleoptera), com
presença de escamas (e.g. Lepidoptera), escopa (e.g. Hymenoptera: abelhas),
antenas filiformes (e.g. Blattaria), pernas raptoriais (e.g. Mantodea), ferrões
alongadas para fazer a postura de ovos em hospedeiros dentro de tecidos
vegetais (e.g Hymenoptera), entre inúmeras outras.
Os insetos ocupam praticamente todos os ambientes
terrestres, com menor abundância e riqueza em regiões polares e marinhas.
Nesses ambientes atuam como detritívoros, saprófagos, herbívoros, predadores,
parasitas, polinizadores, entre outras funções em todos os níveis tróficos. As
ordens que mais se diversificaram foram justamente aquelas relacionadas com as
plantas, especialmente os herbívoros e holometábolos (e.g. Coleoptera e
Lepidoptera, alguns táxons dentro de Diptera e Hymenoptera). Vale abrir um
parêntese e comentar que os insetos podem ser divididos naqueles sem
metamorfose (ametábolos), com metamorfose incompleta (ovo, ninfa e adulto;
hemimetábolos) e os de metamorfose completa (ovo, larva, pupa e adulto;
holometábolos). No caso dos holometábolos, a diferenciação em larva e adulto,
muitas vezes usando recursos alimentares diferentes, possibilitou menor competição
e maiores chances de sobrevivência. Além disso, a especialização em plantas
hospedeiras possibilitou aos insetos a cladogênese paralela a partir da
coevolução com as plantas. Fora os herbívoros, há vários insetos predadores que
regulam as populações de outros insetos (e.g. Mantodea). Este também é o caso
das formigas que predam os herbívoros das plantas que as recompensam com
nectários extraflorais. Há aqueles insetos com adaptações ao hábito
parasitário, com atrofias nos órgãos do sistema digestório (e.g. Phthiraptera)
e perda das asas (e.g. Siphonaptera). Por outro lado, os insetos podem ser
fonte de alimento para diferentes grupos de animais invertebrados (e.g.
aranhas) e vertebrados (e.g. tamanduá; humanos, entomofagia). Eles também
exercem importante influência na ciclagem de nutrientes nos ecossistemas.
A maior parte dos insetos tem hábito solitário, mas
existem aqueles que vivem em sistemas sociais. Exemplos ocorrem em Isoptera e
Hymenoptera, onde há castas distintas, incluindo reprodutiva, divisão de
tarefas, cuidado parental e sobreposição de gerações. Esse tipo de sistema
possibilita maior proteção, obtenção de alimento e, consequentemente,
sobrevivência, embora possa haver também desvantagens (e.g. propagação de
doenças). Porém, os benefícios aparentemente superam os custos na maior parte
dos casos.
A reprodução dos insetos pode se assexuada, como no
sofisticado sistema haplodiplóide das abelhas e formigas, no qual as fêmeas conseguem
gerar novos machos a partir de ovos não fecundados; e fêmeas a partir de ovos
fecundados. Há também outros grupos com diferentes tipos de partenogênese (e.g.
Hemiptera: em afídeos, Thysanoptera, alguns Hymenoptera). Contudo, a maior
parte dos insetos se reproduz sexuadamente, sendo dióicos, com desenvolvimento
direto (e.g. Protura) ou indireto e com várias mudas (e.g. Megaloptera). Geralmente
ocorre cópula e fecundação interna. A maior parte dos insetos é ovípara, mas há
casos de ovoviviparidade (e.g. Hemiptera) e viviparidade (e.g. Diptera: moscas
tsé-tsé).
Os insetos estão incluídos no filo Arthropoda.
Cuvier (1812) delineou a hipótese de que Annelida e Arthropoda formam um táxon
denominado Articulata, baseado em caracteres morfológicos. Entretanto, estudos
mais recentes mostraram que os Annelida poderiam estar mais próximos a táxons
que também possuem larva trocófora (Lophotrochozoa). Hoje, Arthropoda esta no
clado Panarthropoda, sendo grupo irmão de Onychophora, e os dois juntos formando
o grupo irmão de Tardigrada. Quanto aos insetos, foi proposto que eles junto
com Myriapoda formavam o táxon Tracheata, devido à ausência da segunda antena e
presença de túbulos de Malpighi. Mais recentemente, baseado em dados
moleculares e na morfologia do cérebro, foi proposto que os Hexapoda formariam
junto com os Crustacea o táxon Pancrustacea. Dentro dos insetos, há a divisão nos
táxons “Entognatha” e Insecta, os primeiros com aparato bucal internalizado à
cápsula cefálica e os últimos o oposto. Outra divisão forma os Dycondylia, que
possuem dois pontos de articulação na mandíbula. Depois vêm os Pterygota, com a
presença das asas; os Metapterygota, que possuem troncos traqueais, anterior e
posterior, formando um arco nas asas; os Neoptera, com flexão das asas;
Eumetabola, com ausência dos ocelos nos estágios imaturos; e Holometabola (mais
diversos), com metamorfose completa. Finalmente, estudos têm sugerido que tanto
os Arthropoda quanto os Hexapoda são monofiléticos, embora no caso do último,
alguns táxons devam ser reagrupados (e.g. Psocoptera parafilético em relação à
Phthiraptera).
Fonte: maior parte –
Rafael et al. Insetos: diversidade e taxonomia. Ribeirão Preto, Holos
Editora, 2012. 810p.
Poucos detalhes –
Hickman et al. Principios integrados de zoologia. 11ª edição. Rio de
Janeiro: Guanabara Koogan, 2004.
Barnes et al. O invertebrados: uma síntese. São Paulo: Ateneu, 2008
Storer et al. Zoologia Geral. 6ª edição. São Paulo: Companhia Editora,
2003.
5. Morfologia, Diversidade, biologia e
Filogenia dos Aracnídeos
 Escorpião
Escorpião
 Carrapato
Carrapato
 Aranha
Aranha



Os Arachnida (Grego:
arachune, aranha; daqui em diante aracnídeos) são representados por ácaros,
aranhas, escorpiões, pseudoescorpiões, carrapatos e opiliões. Devido a essa
diversidade de formas de vida, alguns autores têm sugerido que os aracnídeos
apresentam maior variedade anatômica que os insetos. Eles são eumetazoários
multicelulares, triploblásticos, protostômios celomados (celoma esquizocélico),
segmentados, com clivagem espiral, desenvolvimento em mosaico e simetria
bilateral.
Os aracnídeos possuem corpo dividido nos tagmas,
prossoma (cefalotórax) e opistossoma (abdômen), com o prossoma geralmente tendo
um par de quelíceras, um par de pedipalpos e quatro pares de apêndices
locomotores. O corpo deles pode variar de alongado, bastante encouraçado, com
pedipalpos raptoriais, segmentação conspícua, com o opistossoma dividido em meso
e metassoma (e.g. escorpiões) até aqueles com formatos esféricos, com
exoesqueleto fino, sem segmentação visível externamente, e com pedipalpos menos
evidentes (e.g. ácaros).
Aracnídeos não possuem mandíbulas. Seu sistema
digestório compreende boca, esôfago delgado ligado a um estômago sugador
operado por músculos que se estendem da sua superfície dorsal até o prossoma,
estômago principal no prossoma com cinco pares de cecos, intestino no
opistossoma, glândula digestiva (fígado), reto, bolsa estercoral e ânus. Suas
peças bucais são geralmente sugadoras ou possuem uma faringe sugadora potente
com a qual ingerem os fluidos e tecidos moles dos corpos de suas presas. Portanto,
percebe-se o sistema digestório modificado para uma alimentação líquida.
A respiração dos aracnídeos se dá pelos pulmões
foliáceos, traquéias ou ambos. Os pulmões foliáceos constituem de uma
evaginação pregueada da parede ventral do abdômen, tendo uma série de lamelas
onde ocorrem as trocas gasosas. Já na
respiração traqueal, o ar entra e sai do corpo desses animas antes/após as
trocas gasosas partir dos espiráculos (orifícios externos situados no prossoma
e opistossoma). Dentro do corpo as traquéias formam “tubos” ramificados que
possibilitam que o ar percorra todo o corpo e ocorram as trocas gasosas. Embora
o último se pareça com o sistema de respiração dos insetos, eles evoluíram de
modo independente.
 Morfologia interna de uma raanha
Morfologia interna de uma raanha

Um dos principais produtos de excreção dos
aracnídeos é a guanina que é removida pelas glândulas coxais, estruturas
saculiformes de parede fina que se abrem nas coxas dos apêndices. Como nos
insetos, as excreções dos aracnídeos também podem ocorrer a partir dos túbulos
de Malpighi, embora esse sistema tenha evoluído independentemente. Nesse caso,
os túbulos trabalham junto com glândulas retais especializas na absorção de
substâncias da hemocele. O potássio juntamente com as excretas são secretados
pelos túbulos, que drenam o fluido (“urina”) para dentro do intestino. Neste
momento, as glândulas retais absorvem a água e o potássio e deixam passar
apenas acido úrico, que é eliminado. Esse sistema conferiu uma excelente vantagem a
esses animais uma vez que a reabsorção de água possibilita habitar ambientes
áridos ou com pequena disponibilidade de água.
O sistema circulatório também é semelhante ao dos
insetos. O coração um tubo delgado, muscular e contrátil, situado dorsalmente
no opistossoma. Ele possui três pares de aberturas (ostíolos) e é circundado
por um pericárdio tubular. Do coração uma aorta caudal estende-se para trás e
uma aorta anterior envia artérias pares para o estômago, pernas, olhos e
glândulas venenosas. O sangue é incolor e contem amebócitos e hemocianina
dissolvida com um pigmento respiratório. O coração bombeia o sangue através das
aortas para dentro dos seios entre os tecidos, quando então passa pelos pulmões
foliáceos para arejamento. O sangue volta pelas por pequenos vasos à cavidade
pericárdica para entrar novamente no coração a partir dos ostíolos.
Os aracnídeos não possuem antena, mas eles possuem
olhos simples com cristalino e retina e algumas espécies, especialmente de
aranhas, enxergam relativamente bem. O sistema nervoso possui um gânglio
bilobado sobre o esôfago, ligado por dois conectivos espessos a uma massa
ganglionar ventral grande, de onde os nervos se irradiam para todos os
órgãos. Além disso, os aracnídeos podem
ter pêlos sensíveis ao tato e diminutos órgãos liriformes situados no corpo e
em suas extremidades, os quais possibilitam a percepção olfativa.
Os aracnídeos também possuem glândulas de seda. Trata-se
de dois ou três pares de fiandeiras em forma de cone, cada uma com um ou muitos
tubos finos através dos quais a seda é secretada por glândulas dentro do
opistossoma para produção de teias, captura de presas, reprodução, abrigo para
a prole e outras funções. No caso da reprodução, os aracnídeos são todos
dioicos e a fecundação é interna. Os ovos são protegidos da dessecação pela
deposição em cavidades úmidas, pela retenção da fêmea (viviparidade) ou pela
presença de um envoltório externo (e.g. seda, folhas dobradas). Há geralmente
rituais complexos de corte (e.g. escorpiões), embora existam casos em que há a
transferência de espermatóforos.
A maioria dos aracnídeos é predadora, se
alimentando de outros artrópodes com especial destaque para os insetos. Para
tal hábito predatório evoluíram nos aracnídeos estruturas inoculadoras
(pedipalpo e quelíceras modificados) e glândulas de veneno. As presas são
agarradas e seguradas pelas quelíceras ou pedipalpos. Então, para imobilizar a
presa o veneno é inoculado e enzimas são secretadas para que a presa seja
reduzida a um liquido ou partículas pequenas que são sugadas. Por outro lado,
seus principais inimigos naturais são as próprias aranhas, aves e alguns
insetos (e.g. vespas parasitoides: Hymenoptera).
Há aproximadamente 70 mil especies de aracnídeos
descritas, sendo aproximadamente a metade delas de aranhas. Eles são
cosmopolitas, com maior diversidade em regiões tropicais. Arachnida pode ser
dividida em 13 ordens (Scorpiones, Uropygi, Schizomida, Amblypygi, Palpigradi,
Araneae, Ricinulei, Pseudoscorpiones, Solpugida, Opiliones, Notostigmata,
Parasitiformes e Acariformes), dependendo da filogenia seguida. Os aracnídeos
são colocados em Arthropoda e dentro do taxon Chelicerata, possuindo como
principais sinapomorfias os quatro pares de pernas e a presença de quilíceras e
pediplapos. Chelicerata seria grupo irmão dos demais Arthropoda (Crustacea,
Hexapoda e Myriapoda). Dentro de Chelicerata parece haver consenso que os
aracnídeos seriam grupo irmão dos extintos Merostomata devido ao prossoma
formar um escudo em forma de carapaça e o primeiro ou segundo metâmero do
opistossoma ser modificado em metâmero genital. De uma maneira geral, parece
haver consenso dos aracnídeos serem monofiléticos.
Fonte: maior parte –
Storer et al. Zoologia Geral. 6ª edição. São Paulo: Companhia Editora,
2003.
Poucos detalhes –
Hickman et al. Principios integrados de zoologia. 11ª edição. Rio de
Janeiro: Guanabara Koogan, 2004.
Barnes et al. O invertebrados: uma síntese. São Paulo: Ateneu, 2008
6. Morfologia, Diversidade, biologia e
Filogenia dos Equinodermos




Estrela do mar

Serpente do mar

Ouriço do mar
Os Echinodermata
(Grego: echinos, espinho; derma, pele; ata, portador de; daqui em diante
equinodermos) são eumetazoários conhecidos como lírios-do-mar (Crinoidea), estrelas-do-mar
(Asteroidea), ofiúros (Ophiuroidea), pepinos-do-mar (Holothuroidea),
ouriços-do-mar e bolachas-da-praia (Echinoidea). Suas principais
características são o endoesqueleto composto por espinhos, sistema
hidrovascular, pedicelárias e brânquias dérmicas. Eles são celomados
enterocélicos, têm ovos com clivagem radial e indeterminada, simetria bilateral
nas larvas (e alguns adultos de Holothuroidea e Echinoidea) e radial pentâmera
nos adultos. Seu corpo pode ser: (1) esférico ou na forma de cálice, preso
aboralmente ao substrato por um pedúnculo e com boca voltada para cima e
circundada por cinco braços (Crinoidea), (2) arredondado e com a superfície
oral em contato com o substrato (Echinoidea), (3) cilíndrico com grande
alongamento no eixo oral-aboral (Holothuroidea), (4) ligeiramente achatado, na
forma de estrela com um disco central que se funde gradualmente com os braços
afilados, e com a boca em posição central na superfície oral (Asteroidea), e
(5) na forma de estrela com braços esguios e abruptamente separados do disco
central (Ophiuroidea). O corpo não é segmentado, possui cinco ou mais áreas
radiais (ambúlacros) que se alternam com áreas interambulacrais. Os
equinodermos possuem corpo composto por tecidos e órgãos.
O endoesqueleto dos equinodermos é formado por
ossículos calcários dérmicos com espinhos, ou por espículas calcárias na derme.
Ele também é coberto por epiderme geralmente ciliada. Em alguns ocorrem as
pedicelárias (exceto Ophiuroidea), que são pequenas mandíbulas acionadas por
músculos e que têm aparência semelhante a pinças. O sistema hidrovascular é de
origem celomática e se projeta da superfície do corpo na forma de uma série de
estruturas tentaculares (pés ambulacrais), protraídos pelo aumento na pressão
do fluido no seu interior. Esse sistema possui uma abertura para o meio externo
(madreporito), que possibilita a saída da água. A locomoção ocorre por meio dos
pés ambulacrais, pelo movimento dos espinhos ou pelo movimento dos braços, que
se estendem a partir do disco central do corpo. O sistema digestório geralmente
é completo, axial ou enrolado, mas com o ânus sendo ausente nos Ophiuroidea. O
celoma é desenvolvido, possui fluido com amebócitos e forma duas cavidades, a
perivisceral e do sistema hidrovascular. Seu sistema circulatório é do tipo
hemal muito reduzido e cercado por extensões do celoma, desempenhando pouca ou
nenhuma função na circulação. Assim, a circulação é feita principalmente pelos
cílios do peritônio. Já a respiração, ocorre a partir de brânquias dérmicas,
pés ambulacrais, árvores respiratórias (Holothuroidea) e bursas (Ophiuroidea).
Os órgãos excretores são ausentes.
Nesses animais não há cabeça ou cérebro, mas um
sistema nervoso constituído por um anel circum-oral e nervos radiais,
geralmente com dois ou três sistemas de redes nervosas localizadas em
diferentes partes do corpo e com complexidade variada, dependendo do grupo. Há
também uma pequena quantidade de órgãos especializados em funções sensoriais
que os possibilitam reconhecerem o ambiente. Por exemplo, receptores táteis e
químicos, pés ambulacrais com ventosas (Ophiuroidea sem ventosas), tentáculos
terminais, fotorreceptores e estatocistos.


Morfologia interna de uma estrela do mar
Os equinodermos são geralmente dioicos, mas algumas
de suas espécies podem ser monóicas. Os sexos são, na maioria das vezes,
externamente iguais. As gônadas são bem desenvolvidas, mas única nos Holothuroidea.
Possuem dutos simples, sem estruturas copulatórias elaboradas ou estruturas
sexuais secundárias. A fertilização ocorre geralmente no meio externo, com
incubação de zigotos em algumas espécies. O desenvolvimento das larvas livre
nadantes pode ser indireto com vários estágios de desenvolvimento ou raramente ocorrer
de modo direto. Podem ocorrer autotomia e regeneração de áreas perdidas. Por
exemplo, algumas espécies de Asteroidea podem regenerar uma estrela
completamente nova a partir de um braço destacado desde que contenha uma parte
do disco central. Portanto pode haver reprodução do tipo assexuada a partir da
clivagem do disco central.
Existem cerca de seis mil espécies de equinodermos
atuais e 20 mil fósseis. Os Ophiuroidea são os equinodermos com maior riqueza
de espécies e provavelmente os mais abundantes. Os equinodermos possuem cores
crípticas ou vivas, que podem variar de amarelo, vermelho, branco, verde ou parda
(e.g. Crinoidea). Eles podem variar em tamanho de um centímetro a um metro. Todos
vivem em ambiente marinho e em diferentes profundidades. Algumas especies são
pelágicas, mas a maior parte é bentônica. Nesses ambientes os equinodermos podem
ser predadores (e.g. Asteroidea), necrófagos (e.g. Ophiuroidea), filtradores
(e.g. Ophiuroidea), detritívoros (e.g. Holothuroidea) e suspensívoros (e.g. Crinoidea).
Por outro lado, diferentes animais como, ctenóforos, turbelários, cirripédios,
copépodes, gastrópodes, bivalves e peixes, se estabelecem no interior ou sobre
os equinodermos, ou ainda podem predá-los. Os Asteroidea passam a maior parte
de seu tempo presos a algum substrato sólido (e.g. rochas), embora algumas
possam viver sobre areia ou lodo. Superfícies ásperas e verticais facilitam com
que o animal se prenda e locomova-se com os pés ambulacrais portadores de
ventosas. Já os Asteroidea de ambientes arenosos/lodosos não têm ventosas e
seus pés ambulacrais pontiagudos, os quais são inseridos no substrato para dar
apoio para que o animal se desloque a frente. Esses animais se alimentam de
moluscos, crustáceos, anelídeos, entre outros.
Os Ophiuroidea têm o comportamento de se esconder
embaixo de pedras, plantas marinhas ou enterrando-se no lodo ou areia durante o
dia. À noite eles são mais ativos, quando se movem-se por movimento
serpentiformes rápidos possibilitados por segurarem-se a objetos com alguns
braços e sendo impulsionados por outros. Eles tambem podem nadar. Alimentam-se
de pequeno crustáceos, moluscos e outros animais ou detritos associados ao
fundo. Por outro lado, são predados por peixes.
Os Echinoidea vivem em rochas ou lodo nas praias e
no fundo do mar. Locomovem-se pelo uso simultâneo dos espinhos e pés
ambulacrais. Os pés servem apara agarrar objetos no fundo do mar, assim como
para segurar pedaços de detritos contra o corpo do animal para camuflá-lo.
Todos eles limpam seu corpo com o movimento dos espinhos e pedicelárias. As
dejeções do ânus são removidas de modo semelhante. A maior parte dos ouriços se
alimenta de plantas marinhas, matéria animal morta e pequenos organismo
raspados pela lanterna de Aristóteles, uma estrutura complexa de cinco lados e
cinco dentes inserida dentro da boca desses animais. Já as bolachas-da-praia
alimentam-se de partículas orgânicas da areia ou do lodo por meio da ingestão
direta do substrato ou por meio das redes de muco. Os Echinoidea têm muitos
ciliados comensais no sistema digestório, mas tambem comensais externos e
parasitas internos e externos em seu corpo. Peixes, estrelas-do-mar, caranguejos,
aves e mamíferos predadores são seus principais inimigos naturais.
Os Holothuroidea movem-se lentamente no fundo do
mar a partir de seus pés ambulacrais ou movimentos musculares do corpo. Eles
também cavam no lodo ou areia da superfície, onde se enterram e deixam somente
as extremidades do corpo expostas. Quando perturbadas contraem-se lentamente. A
alimentação é constituída principalmente por matéria orgânica dos detritos do
fundo, a qual é empurrada para a boca. Outro tipo de alimento é o plâncton que
fica aprisionado no muco de seus tentáculos. Vários comensais e parasitos vivem
sobre ou dentro dos Holothuroidea, incluindo anelídeos, caranguejos e peixes
(e.g. Carapus spp.). Alguns Holothuroidea
possuem órgãos de Cuvier, que é associado ao reto e usado como defesa contra
predadores. Nesse caso, quando perturbados esses Holothuroidea lançam esses
órgãos para fora do ânus para imobilizar o atacante com uma massa pegajosa de
tubos adesivos. Há também espécies que quando ameaçadas rompem a parede do corpo
e eliminam diversos órgãos, os quais são regenerados depois.
Já os Crinoidea são, em sua maioria, sedentários
vivendo desde abaixo da linha de maré até profundidades abissais. Entretanto,
há espécies de vida livre como as do gênero Antedon,
que nadam usando seus braços longos e agarrando-se a objetos no fundo com seus
cirros. Seu principal alimento é o plâncton microscópico e detritos, os quais
são colhidos pelos tentáculos e dirigidos pelos cílios à boca. Esses animais
põem os ovos na água ou os ovos permanecem presos em seu corpo (pínulas) até a
eclosão. Assim como outros equinodermos, os Crinoidea têm grande capacidade
regenerativa, soltando os braços ou grande parte do cálice para, então, renovar
essas partes. Há muitos comensais e parasitos, com destaque para os poliquetos
(e.g. Myzostomum). Alguns pequenos
gastrópodes perfuram os Crinoidea para se alimentar de suas partes moles.
Os
equinodermos são animais pertencentes ao táxon Deuterostomia. Entretanto, a
filogenia deles ainda é controversa. Se levado em conta as larvas bilaterais e
o esqueleto interno bem desenvolvido dos equinodermos, pode-se sugerir que eles
têm um ancestral em comum com os Hemichordata e formam o clado Ambulacraria (larva
dipleurula). Porem, a divergência anatômica entre os adultos desses dois filos
e o longo registro fóssil de ambos sugerem que eles evoluíram ao longo de
linhas separadas durante um longo período de tempo. Dentro de Echinodermata,
tem-se sugerido que os Crinoidea são o grupo mais antigo e mais próximo de
táxons extintos (Homalozoa e Helicoplacoroidea). Asteroidea e Ophiuroidea
parecem mais semelhantes entre si, embora as formas larvais sejam diferentes,
assemelhando-se larvas de Ophiuroidea às de Echinoidea. Também, os
Holothuroidea e Asteroidea parecem muito diferentes entre os adultos, mas as
larvas são estruturalmente semelhantes. Portanto, uma inferência clara ainda
não é possível, mas autores têm produzido cladogramas baseados em politomias
seguindo a escala evolutiva de Crinoidea, Asteroidea, Ophiuroidea,
Echinodermata e Holothuroidea, nessa sequência, respetivamente.
Fonte: maior parte –
Hickman et al. Principios integrados de zoologia. 11ª edição. Rio de
Janeiro: Guanabara Koogan, 2004.
Storer et al. Zoologia Geral. 6ª edição. São Paulo: Companhia Editora,
2003.
Poucos detalhes –
Barnes et al. O invertebrados: uma síntese. São Paulo: Ateneu, 2008
7. Evolução, biologia e diversidade dos Peixes


“Peixes” é o nome dado a um grupo de animais que
vivem no ambiente aquático, que possuem brânquias, membros modificados em
nadadeiras (se presentes) e escamas de origem dérmica no tegumento (atuais:
Myxinoidea, Petromyzontoidea, “Agnatha” e parte dos Gnathostomata). O termo peixes
é usado por conveniência e reflete um táxon parafilético, uma vez que exclui os
Tetrapoda. A evolução dos peixes começou há cerca de 530 milhões de anos, na
explosão de diversidade do período Cambriano.
Foi nesse período que os primeiros cordados desenvolveram o crânio e
coluna vertebral, dando origem aos primeiros craniados e vertebrados, que são
os peixes.
O ancestral dos
peixes seria um protocordado livre nadante do início do Cambriano, com o qual possivelmente
compartilharam a notocorda, tubo nervoso dorsal, bolsas faríngeas e cauda pós
anal. Os primeiros peixes representariam um conjunto de táxons parafiléticos sem
maxilas (agnatos), cujo registro mais antigo é dos Haikouichthys. No final do Cambriano surgiram os conodontes e os
pequenos peixes com corpo constituído por placas de armadura, os ostracodermes.
Os agnatos atuais incluem as feiticeiras (Myxinoidea) e lampreias (Petromyzontoidea).
A ancestralidade das lampreias e feiticeiras é incerta, pois há pouca
semelhança entre si e também com os ostracodermes. Feiticeiras e lampreias,
embora não possuam vértebras ou elas sejam rudimentares, ainda assim são
inseridas em Vertebrata por possuírem crânio e outras homologias de
vertebrados. Dos agnatos, se diferenciou uma linhagem dos peixes gnatostomados
(com maxilas). Portanto, todos os demais peixes com exceção as atuais lampreias
e feiticeiras possuem apêndices pareados e maxilas, e são incluídos juntamente
com os tetrápodes, formando a linhagem monofilética dos gnastotomados. Esses
apareceram no registro fóssil do final do Siluriano com as maxilas totalmente
formadas. Sendo assim, não há formas intermediárias conhecidas entre ágnatos e
gnatostomados.
No Devoniano houve grande diversificação dos peixes
com maxilas como os placodermos, os quais evoluíram dos ostracodermes e, assim
como eles, possuíam pesada armadura protetiva no corpo. Outra linhagem que
evoluiu no mesmo período foi a dos Acanthodii, os peixes de nadadeira com
espinhos, que são colocados como grupo irmão dos peixes ósseos (Osteichthyes).
Os placodermos se tornaram extintos no Carbonífero, já os Acanthodii foram
extintos no Permiano inferior. Mas, antes, no final do Siluriano também surgiu
a linhagem dos peixes cartilaginosos (Chondrichthyes), os quais perderam a
armadura pesada e desenvolveram um esqueleto cartilaginoso no lugar de ósseo.
Esse grupo formou duas linhagens, a dos elasmobrânquios (tubarões e raias) e
holocéfalos (quimeras). Ambas as linhagens se diversificaram muito nos períodos
Devoniano e Carbonífero, mas também quase se extinguiram no final do Permiano
(final da era Paleozóica). Elas se recuperaram no início da era Mesozóica e, em
especial os elasmobrânquios, se tornaram um grupo bem estabelecido nos mares,
onde estão presentes até hoje (Cenozóico) como, principalmente, predadores
ativos. Tal diversidade desses animais é relacionada à presença de maxilas com
dentes, as quais proporcionaram mordedura mais forte e, consequentemente, maior
facilidade de obter alimento.
Por sua vez, os peixes ósseos se diversificaram em
duas linhagens distintas, os peixes com nadadeiras raiadas (Actinopteygii), que
se irradiaram e deram origem aos peixes atuais, e os peixes com nadadeiras lobadas
(Sarcopterygii), os quais são menos diversificados, mas representam o grupo
irmão dos tetrápodes. A linhagem dos Actinopteygii se diversificou bastante no Siluriano
e Devoniano, mas no fim do Permiano houve uma drástica redução. Na era
Mesozóica, a diversificação votou a acontecer com crescente aumento até a atual
era Cenozóica. Essa diversificação se tornou possível a partir das varias
adaptações desenvolvidas por esse grupo como a presença dos opérculos sobre as
brânquias (eficiência respiratória), bexiga natatória (flutuabilidade) e a
musculatura das maxilas, que possibilitou a diversificação dos hábitos
alimentares, entre outras. Já a os Sarcopterygii se diversificaram bastante no
Devoniano e Carbonífero, com gigantesca redução a partir do Carbonífero, quando
então se mantiveram como um relictual grupo.
Entre os agnatos atuais, as feiticeiras constituem
um grupo exclusivamente marinho, encontrado especialmente no Atlântico Norte e
que possui o hábito parasitário ou predatório. Feiticeiras se alimentam
principalmente de anelídeos, crustáceos, moluscos e peixes mortos ou
moribundos. São animais praticamente cegos, mas que desenvolveram um sensível
olfato que capta o cheiro de animais mortos ou moribundos. Então, as
feiticeiras penetram no corpo do animal por um de seus orifícios ou o escavam.
Depois, elas raspam o corpo da presa/hospedeiro e retiram pedaços de carne para
se alimentar. Para fazer isso, elas possuem duas placas de dentes sobre a
língua e que se movem funcionando com um torquês. Para se pressionar e se
prender firmemente ao corpo das presas, as feiticeiras torcem seu corpo
formando um nó em sua cauda. Por outro lado, quando atacadas as feiticeiras
secretam um muco leitoso e branco, o qual é expelido a partir de glândulas
especiais posicionadas ao longo do corpo para se defenderem. Esse muco é tão
escorregadio que se torna difícil segurar o animal. A biologia reprodutiva das
feiticeiras é pouco conhecida, mas já se sabe que a razão sexual é desviada
para as fêmeas (100x1), que elas produzem poucos e grandes óvulos de dois a sete
centímetros de comprimento, com muito vitelo e não há larvas.
As lampreias são todas registradas para o
hemisfério Norte e podem atingir um metro de comprimento. A maior parte de suas
espécies é parasita e utiliza a boca em forma de ventosa e com pequenos
dentículos para se aderir ao corpo do hospedeiro. As lampreias sugam fluidos
corporais dos hospedeiros e para manter a circulação ativas, elas injetam
anticoagulantes nas feridas que desferem em seus hospedeiros. Lampreias não
parasitas não se alimentam quando adultas, vivendo tempo suficiente apenas para
reprodução. Todas sobem aos rios para se reproduzirem (anádromas). Os machos constroem os ninhos no fundo dos
rios e quando as fêmeas chegam, eles as agarram pelo lado dorsal da cabeça,
esperam elas depositarem os óvulos e em seguida os fertilizam. Os ovos são
pegajosos e se aderem a pedregulhos, sendo posteriormente cobertos por areia. As
larvas eclodem em duas semanas, passam por um estágio filtrador (amocete) até se
metamorfosear e tornar adulta.
Os peixes com esqueleto cartilaginoso são
encontrados em ambientes marinhos e dulciaquícolas. Os tubarões possuem corpo fusiforme
e hidrodinâmico dotado de escamas ásperas (placóides), maxilas com muitos
dentes em fileiras de reposição, com órgãos sensoriais (e.g. ampola de
Lorenzini: eletrorreceptor) e linha lateral com neuromastos, todos aguçados
para uma vida ativa com hábito predatório, embora existam espécies filtradoras
(e.g. Cetorhinus maximus). Alimentam-se
de diferentes animais marinhos como peixes, especialmente em cardumes, e
mamíferos, sendo predadores de topo destes ambientes. Seus inimigos naturais
geralmente são outros tubarões. As raias possuem corpo achatado, ferrão com
espinhos, às vezes com veneno (defesa) e com nadadeiras peitorais muito
alargadas adaptadas para uma vida bentônica, também com hábito predatório. As
raias possuem dentes adaptados para triturar ou órgãos elétricos para atordoar suas
presas, sendo elas principalmente moluscos, crustáceos e pequenos peixes. Já as
quimeras possuem maxilas com placas achatadas, sendo a maxila superior fundida
ao crânio (exclusividade). Sua alimentação consiste em algas marinhas,
moluscos, equinodermos, crustáceos e peixes. Todos os peixes cartilaginosos são
dioicos, com fecundação interna, com oviparidade, ovoviviparidade e
viviparidade, e desenvolvimento direto. Os tubarões e raias possuem clásper, um
órgão copulatório intromitente formado da modificação na nadadeira pélvica.


Morfologia interna peixes
Os peixes ósseos ocupam todos os ambientes aquáticos do planeta, de mares polares até o Equador, e da superfície até profundidades superiores a 9000 metros. Vivem em águas abertas, fundo arenoso, rochoso e lodoso, em fendas dos recifes de corais, estuários, rios e lagos de água doce ou alcalina, em cavernas e em fontes termais. A maior parte é pelágica e carnívora, mas há onívoros, herbívoros, parasitas, planctófagos, comensais e saprófagos. A posição da boca diz muito quanto ao hábito alimentar e ambiente em que vivem. Por exemplo, boca voltada para baixo (posição ventral) geralmente pode ser relacionada ao hábito saprófago e associado ao fundo (e.g. cascudos, família Loricariidae). Os inimigos naturais dos peixes ósseos são os mais diversos animais, muitas vezes sendo outros peixes ósseos. A maior parte dos peixes ósseos é dioica, com fecundação externa e ovípara, embora existam ovovivíparos (e.g. Hypsurus) e vivíparos (e.g. Zoarces viviparus). Há aqueles com cuidado parental, que protegem os ovos na boca (e.g. Opistognathus macrognathus), aqueles cujos machos possuem bolsa no corpo e encubam os ovos (e.g. Hippocampus spp.), entre outras das mais variadas estratégias reprodutivas, variando de semélparos a iteróparos. Vale ressaltar que, ainda, pode haver mudança de sexo e com produção de gametas relacionados a estes ao longo da vida de algumas espécies de peixe. Alguns peixes ósseos fazem migrações, do oceano para os rios (anádromos) ou vice-versa (catádromos) para se reproduzir e/ou alimentar.
Os peixes
possuem mais de 25 mil espécies descritas, tendo maior riqueza e diversidade
que todos os demais vertebrados juntos (todos os vertebrados: ~48 mil
espécies). Destas, aproximadamente 90 são agnatos (lampreias e feiticeiras), 850
são cartilaginosos e mais que 24 mil são peixes ósseos. O corpo deles varia muito
em formas e cores, mas podem-se citar as formas longas, delgadas cilíndricas e
com a cauda comprimida (e.g. lampreias e feiticeiras), fusiforme (e.g. tubarões
e peixes ósseos como Micropogonias furnieri),
achatado dorso-ventralmente (e.g. raias e diversos peixes ósseos, como: peixe-morcego Ocgocephalus vespertilio), achatado
lateralmente (e.g. paru: Pomacanthus paru),
globular (e.g. baiacu: Cyclichthys
spinosus), anguiliforme (e.g. peixe-espada Trichurus
lepturus), e de cores crípticas, vistosas a iridescentes.
Fonte: maior parte –
Hickman et al. Principios integrados de zoologia. 11ª edição. Rio de
Janeiro: Guanabara Koogan, 2004.
Storer et al. Zoologia Geral. 6ª edição. São Paulo: Companhia Editora,
2003.
Poucos detalhes –
Bemvenuti e Fischer. 2010. Peixes: morfologia e adaptações. Cadernos de Ecologia
Aquática 5 (2):31-54.
Pough et al. A vida dos vertebrados. 4ª edição. São Paulo: Atheneu
Editora, 2008.
8. Evolução, biologia e diversidade dos
Anfíbios e Répteis






Cobra

Crocodiliano

Lagarto

perereca

cecília
Os anfíbios e os
répteis pertencem ao táxon Tetrapoda e evoluíram de um Sarcopterygii ancestral,
ou seja, um peixe de nadadeiras lobadas e com pulmão. Toda esta história
evolutiva aparenta ter começado no período Devoniano (~400 m.a.a.), o qual foi
muito instável com alternância em épocas de seca e de inundação. Nas épocas de
seca, as águas dos rios e lagos eram disponíveis em pequenas poças e com pouco
oxigênio dissolvido. Sendo assim, os peixes de água doce que conseguissem se
deslocar entre as poças e retirar o oxigênio do ar atmosférico seriam capazes
de sobreviver e se diversificar. Foi nesse período que alguns vertebrados
aquáticos, os sarcopterígios, já tinham pulmões e desenvolveram membros
locomotores, os quais representaram a base para a evolução dos tetrápodes (incluindo
anfíbios e répteis) e sua conquista do ambiente terrestre. Tanto é que apenas
os peixes com nadadeiras lobadas e pulmão desenvolvido como um divertículo da
faringe sobreviverem em ambientes de água doce nesse período. Esses peixes, ao
longo dos milhares de anos, desenvolveram um sistema de circulação dupla
(circulação sistêmica e pulmonar) e membros articulados, todas essas
características em conjunto sendo a base para a evolução do ancestral dos tetrápodes.
Eusthenopteron foi um peixe de nadadeiras
lobadas e pulmonado do devoniano que possuía ossos como úmero, rádio e ulna,
homólogos aos ossos do pulso dos Tetrapoda. Ele provavelmente conseguia saltar
entre poças de lodo. No final do Devoniano surgiram os Acanthostega, um dos primeiros tetrápodes com membros locomotores bem
formados e com dígitos distintos. Nesse mesmo período surgiram os Ichthyostega, os quais possuíam cintura
escapular desenvolvida, membros com ossos robustos e, provavelmente, já
conseguiam se arrastar na terra. Assim, esses dois últimos exemplos citados
representam os primeiros tetrápodes com adaptações ao ambiente terrestre.
Já no período Carbonífero, quando a Terra teve um
clima quente e úmido, com florestas de musgos e pteridófitas, e grande
abundância de insetos, os Tetrapoda se irradiaram rapidamente e originaram os
Lissamphibia. Os anfíbios atuais se diversificaram de uma linhagem desses
Lissamphibia no início da era Mesozóica. Eles se dividiram em três linhagens:
as cecílias (Gymnophiona), salamandras (Urodela) e sapos, pererecas e rãs (Anura).
Hoje todos eles são inseridos em um grupo denominado temnospôndilos, devido aos
quaro dígitos nos membros anteriores ao invés dos cinco da maioria dos outros
tetrápodes.
Já os répteis e demais amniota se diversificaram no
final do Carbonífero com aumento em diversidade no final do Permiano. Eles se
originaram a partir de tetrápodes semelhantes aos anfíbios, os antracosauros,
com os quais compartilharam modificações de ossos do crânio e cinco dígitos. Ao
final do Carbonífero os amniota se dividiram em três linhagens, baseadas na
morfologia do crânio: os anápsidos (tartarugas), diápsidos (tuataras, lagartos,
serpentes, anfisbenas, crocodilianos e aves) e sinápsidos (mamíferos). Destes, os diápsidos se dividiram em
lepidossauros (tuataras, lagartos, serpentes, anfisbenas), arcossauros
(crocodilianos e aves) e sauropterígios (extintos, como plesiossauros). Como os
répteis possuem um ancestral em comum com as aves, seu táxon, Reptilia,
representa um grupo parafilético. Assim, é correto dizer que os répteis são
apenas amniotas que não são aves.
Como o nome diz anfíbios são animais que possuem
uma vida dupla, com fase larval geralmente associada ao ambiente aquático e
fase adulta ao terrestre, assim, são animais semiterrestres. São comuns em
regiões temperadas úmidas, mas a maior parte é tropical. Eles são os primeiros
cordados a viverem fora da água e os vertebrados terrestres mais abundantes em
muitas florestas tropicais. Alguns poucos também podem viver em desertos, onde
se escondem em buracos durante o dia e são ativos à noite. Há alguns anfíbios
que são exclusivamente aquáticos, especialmente algumas salamandras (e.g. Cryptobranchus spp., Necturus spp., Amphiuma spp.). Outros são arborícolas, se escondem em baixo de
troncos ou pedras e ainda outros se enterram na terra úmida, podendo haver
migração entre esses ambientes. Eles são importantes predadores de insetos e de
outros animais invertebrados. São importantes animais na ciclagem de nutrientes
entre a água doce e os ambientes terrestres, pois os nutrientes ingeridos por
suas larvas em ambiente aquático são transportados para a terra através da
dispersão e morte dos adultos metamorfoseados. Por outro lado, servem como
alimento para diversos vertebrados, inclusive humanos. Alguns anfíbios como os
sapos tambem possuem cordas vocais na laringe que os possibilitam vocalizar e
chamar parceiros para o acasalamento. A vocalização também pode ser usada como
caráter taxonômico, distinguindo espécies crípticas. São animais dioicos, com
os machos geralmente tendo músculos dos braços mais fortes e dedos mais curtos
que possibilitam o amplexo. A reprodução geralmente tem corte e ocorre na água
e com fertilização externa. Os ovos são envolvidos em camadas gelatinosas e as
larvas têm desenvolvimento indireto e dentro da água. Há espécies de
salamandras em que ocorre a transferência de espermatóforos. Outras são
pedomórficas, ou seja, os adultos maduros sexualmente ainda possuem
características larvais como a permanência das brânquias (e.g. Axolote).
Finalmente, há em muitos casos, cuidado parental, com o anfíbio protegendo seus
ovos no dorso, em folhas ou até dentro da boca.



Morfologia de um anfíbio

Reprodução de um anfíbio
Os répteis foram impressionantemente abundantes durante
um período de aproximadamente 165 milhões de anos (parte da era Mesozóica), no
qual os dinossauros foram extremamente dominantes no ambiente terrestre e
aquático. Devido à extinção em massa do final da era Mesozóica, sobreviveram poucas
linhagens atuais. Entre os sobreviventes, a tuatara é o único representante de
um dos mais antigos grupos, cujos outros representantes desapareceram há 100
milhões de anos (Sphenodon). A maioria dos répteis vive em regiões tropicais e
subtropicais, ocupando uma ampla variedade de habitats como fendas de rochas,
árvores, casas, buracos de outros animais, desertos, e alguns se enterram na
areia. Alguns, como as tartarugas, vivem dentro ou perto da água. São animais
ectotérmicos, assim como os anfíbios, o que faz com que dependam da temperatura
do ambiente para desenvolverem suas atividades.
A maioria dos répteis é carnívora, embora existam jabutis e algumas
tartarugas que se alimentam de tecidos vegetais. A iguana-de-crista (Amblyrhynchus cristatus) mergulha no mar
e nada para lugares onde possa pastar algas marinhas. Lagartos e pequenas
cobras se alimentam de insetos e outros invertebrados; pequenas tartarugas se
alimentam de cnidários e outros invertebrados marinhos; grandes lagartos,
cobras e crocodilianos comem vários vertebrados, de peixes a mamíferos. Muitos
répteis predadores percebem suas presas a partir da visão e o movimento da
presa. Ainda, nos diápsidos se desenvolveu o crânio cinético com a presença do
osso quadrado, os quais possibilitam maior abertura da boca e,
consequentemente, a captura de presas maiores. Muitas cobras inoculam venenos
em suas presas a partir de sua dentição modificada para tal (e.g. solenóglifa),
enquanto outras constringem suas presas matando-as por asfixia. Alguns jabutis
e tartarugas podem viver até 100 anos e alguns crocodilos e cobras de 25 a 40
anos. A reprodução desses animais geralmente envolvem rituais de corte e
cópula. A fecundação é sempre interna e a maioria é ovípara. Há também cobras e
lagartos ovovivíparos e vivíparos. Os ovos possuem cascas endurecidas por sais
calcários nos crocodilianos, algumas tartarugas e lagartixas. A forma geral dos
ovos é oval longa, embora possa ser esférica nos jabutis e tartarugas. O número
de ovos produzidos pode variar muito entre as espécies. Por exemplo, algumas
tartarugas marinhas (e.g. Caretta
spp.) podem produzir anualmente 400 ovos, mas há algumas lagartixas que
produzem um único ovo nesse mesmo intervalo de tempo. Em cobras vivíparas já
houve o registro do nascimento de 70 filhotes (Thamnophis sp.). Todos os embriões são circundados pelas membranas
do âmnio, cório e alantóide, as quais o protegem contra a dessecação e choque
físico durante o desenvolvimento. O desenvolvimento é direto e, ao eclodir, o
jovem é independente.

Há aproximadamente 4300 espécies de anfíbios, das quais 160 são cecílias, 360 são salamandras e mais que 3500 são sapos, pererecas e rãs. Eles variam de poucos centímetros a mais de um metro e meio, de cores crípticas a apostemáticas, de organismos com corpo sem apêndices, alongado e de hábito fossorial (cecílias), alongadado e com cauda (salamandras), e robusto e adaptado ao salto, natação ou aderência em superfícies sólidas (sapos, pererecas e rãs). Os répteis possuem uma diversidade ainda muito maior em riqueza e formas que os anfíbios. Em riqueza, são descritas aproximadamente 7000 espécies. Em forma, variam daqueles com corpo arredondado com presença de carapaça, plastão e com fusão de costelas e vértebras (tartarugas), corpo alongado e membros reduzidos (lagartixas), corpo cilíndrico e ausência de membros (anfisbenas), corpo alongado com vértebras curtas e largas e ausência de membros (serpentes), e corpo e crânio alongados e robustos (crocodilianos). Elas podem ter de poucos centímetros aos incríveis sete metros de comprimento (Crocodylus porosus). As cores tambem variam muito de crípticas, a vivas e apostemáticas.

Morfologia de um réptil
Há aproximadamente 4300 espécies de anfíbios, das quais 160 são cecílias, 360 são salamandras e mais que 3500 são sapos, pererecas e rãs. Eles variam de poucos centímetros a mais de um metro e meio, de cores crípticas a apostemáticas, de organismos com corpo sem apêndices, alongado e de hábito fossorial (cecílias), alongadado e com cauda (salamandras), e robusto e adaptado ao salto, natação ou aderência em superfícies sólidas (sapos, pererecas e rãs). Os répteis possuem uma diversidade ainda muito maior em riqueza e formas que os anfíbios. Em riqueza, são descritas aproximadamente 7000 espécies. Em forma, variam daqueles com corpo arredondado com presença de carapaça, plastão e com fusão de costelas e vértebras (tartarugas), corpo alongado e membros reduzidos (lagartixas), corpo cilíndrico e ausência de membros (anfisbenas), corpo alongado com vértebras curtas e largas e ausência de membros (serpentes), e corpo e crânio alongados e robustos (crocodilianos). Elas podem ter de poucos centímetros aos incríveis sete metros de comprimento (Crocodylus porosus). As cores tambem variam muito de crípticas, a vivas e apostemáticas.
Fonte: maior parte –
Hickman et al. Principios integrados de zoologia. 11ª edição. Rio de
Janeiro: Guanabara Koogan, 2004.
Storer et al. Zoologia Geral. 6ª edição. São Paulo: Companhia Editora,
2003.
Poucos detalhes –
Pough et al. A vida dos vertebrados. 4ª edição. São Paulo: Atheneu
Editora, 2008.
9. Evolução, biologia e diversidade das
Aves





Reconstituição de Archaeopteryx

Registro fossil Archaeoptery

Tucano

Seriema
Segundo hipóteses tradicionais, a mais antiga “ave”
conhecida, Archaeopteryx lithographica,
viveu no período Jurássico Superior há cerca de 147 milhões de anos (Era Mesozóica).
Essa “ave” possuía penas como as atuais, porém com bico curto e pequenos dentes
ósseos implantados em alvéolos, longa cauda óssea, dedos com unhas e costelas
abdominais. Tendo em vista as características típicas reptilianas (e.g.
esqueleto, dedos com unhas, etc.), tem-se sugerido que as aves se derivaram dos
dinossauros, especificamente dos Theropoda, os quais possuíram pelo menos cinco
linhagens com estruturas semelhantes a penas no corpo (e.g. coelosaurus e
dromeossauros). Entretanto, estudos mais recentes sugerem que Archaeopteryx não é uma ave, sendo
inserida no clado dos dinossauros não avianos: Deinonychosauria. Mesmo assim, é
possível que as aves tenham derivado desse mesmo grupo, sendo as primeiras aves
possivelmente pertencentes aos gêneros Jeholornis
e Sapeornis, pouco conhecidos. Portanto,
as aves são consideradas atualmente como dinossauros terópodos derivados, compartilhando
com os demais terópodos o marcante pescoço alongado e móvel em forma de “S”,
entre outras.
A evolução dos Theropoda e, especificamente aves, sugere
que as penas surgiram antes do voo. Dinossauros não avianos provavelmente
utilizavam as penas em combinação com posturas do corpo para se interagir intra
ou interespecificamente ou para cobrir os ovos nos ninhos. Alguns Theropoda,
como os dromeosaurus, eram bípedes e possuíam estruturas no punho que os
possibilitavam flexioná-los lateralmente enquanto os movimentavam em giros. Tanto
as penas quanto o caminhar bípede, o punho flexível e a modificação dos membros
anteriores em asas foram importantes para voo. Mas, antes do voo, as asas
teriam evoluído e possibilitado a captura de presas (hipótese “do chão para
cima”). Neste caso, os Archaeopteryx e
outros maniraptores, já alados, corriam atrás de suas presas voadoras (insetos),
saltavam e estendiam suas asas para derrubar as presas no chão e facilitar a
apreensão delas pelas suas garras e dentes. Então, os complexos movimentos
feitos pelas asas e o pulso flexível passaram a ser utilizados para dar propulsão
e equilíbrio aos saltos, bem como capturar os insetos no ar. Tais
características possibilitaram o hábito predatório dos maniraptores (especialmente
dromeosauros), tornando-os um dos mais eficientes grupos de predadores
terrestres da Era Mesozóica. Depois do salto e planagem quando correndo atrás
de suas presas, o voo teria surgido nas aves primitivas (Avialae) a partir da
corrida e decolagem batendo as asas.
Nas aves atuais (Neornithes), o corpo aerodinâmico
com ossos pneumáticos, a quilha, onde a musculatura se insere, fúrcula, as
penas com raque, vexilo e cálamo e tendões possibilitaram o voo mais ativo (batido)
e o empoleiramento. Outras aves, as que não voam (ratitas), desenvolveram corpo
largo (estabilidade na água), plumagem densa (flutuabilidade e isolamento
térmico), glândula uropigial (impermeabilização), membranas interdigitais (nado),
entre outras. Tais características representaram e, ainda representam, uma vantagem
no escape de predadores, na captura de presa, evitação de competição, na
dispersão de ambientes inóspitos, entre outras. Tendo em vista essas
adaptações, no Cretáceo (Era Mesozóica) e no início do Terciário (Era
Cenozóica) ocorreu a irradiação das aves, resultando nas 27 ordens atualmente
conhecidas e o segundo maior grupo de vertebrados (perdendo em riqueza apenas
para os peixes).
Hoje são descritas mais de 11200 espécies de aves,
distribuídas por quase toda a Terra, sendo aproximadamente 1800 espécies no
Brasil. Elas ocorrem em florestas, desertos, montanhas, pradarias, cavernas,
oceanos e algumas poucas até visitam os Polos Norte e Sul. Há aquelas
corredoras terrestres (ratitas: e.g. avestruz, ema e casuar), as
tradicionalmente voadoras (maior parte dos Neognathae) e as aquáticas (e.g. Sphenisciformes:
pinguins). Sua diversidade morfológica é restrita, uma vez que possuem corpo em
uniformidade estrutural adaptada para o voo. Quanto a cores, elas são muito
diversificadas, de crípticas (geralmente associadas ao pigmento melanina) a
coloridos vistosos (feomelaninas e carotenóides) ou iridescentes (principalmente
melanina). Já quanto à vocalização, são muito diversas, sendo os cantos muitas
vezes utilizados para distinguir as espécies regionalmente. O canto serve para
comunicação intra e interespecífica em interações envolvendo territorialidade,
reprodução e migração e defesa.
As aves são animais endotérmicos (regulam sua
temperatura corpórea) e, portanto, são ativas em todas as estações do ano. Elas
podem migrar quando as condições ambientais são desfavoráveis ou para se
reproduzir. Seus hábitos alimentares são dos mais variados e a morfologia dos
bicos ajuda a compreendê-los. Por exemplo, aves com bicos robustos e
pontiagudos nas pontas, como dos Falconidae (e.g. caracarás), são geralmente predadoras,
usando o bico para dilacerar suas presas (e.g. répteis, outras aves e pequenos mamíferos).
Há também aves insetívoras (pica-pau e curiango), filtradoras (flamingo), frugívoras/granívoras
(tentilhão), piscívoras (garça), saprófagas (urubu) e nectarívoras (beija-flor),
entre outras.


Morfologia externa das aves

Morfologia interna das aves
Fonte: maior parte –
Pough et al.
A vida dos vertebrados. 4ª edição. São Paulo: Atheneu Editora, 2008.
Poucos detalhes –
Hickman et al. Principios integrados de zoologia. 11ª edição. Rio de
Janeiro: Guanabara Koogan, 2004.
Storer et al. Zoologia Geral. 6ª edição. São Paulo: Companhia Editora,
2003.
10. Evolução, biologia e diversidade dos
Mamíferos






Panda

Leão

Anta

Tamanduá bandeira

Porco
Os mamíferos
evoluíram de animais amniotas que possuíam uma fenestra temporal mais baixa na
cabeça, os sinapsídeos. Esses últimos foram o grupo mais dominante de amniotas
do final da Era Paleozoica (~290 m.a.) e de toda a Era Mesozóica (~225-65 m.a.),
sendo considerados os carnívoros de topo do período Permiano ao Triássico. Contudo,
havia sinapsídeos herbívoros, como é caso dos pelicossauros que viveram no
inicio do Permiano (~290 m.a.). Outra linhagem de sinapsídeos era a dos
terápsidos, os únicos carnívoros que sobreviveram após a Era Paleozóica. Nesses
animais ocorreu pela primeira vez o andar ereto sobre quatro patas, o que
reduziu a estabilidade do corpo com o solo, mas também ocorreu a expansão do
cerebelo, o centro de coordenação muscular do encéfalo (o início para o
desenvolvimento de um sistema nervoso altamente desenvolvido nos mamíferos).
Então, os terápsidos irradiaram-se e deram origem a várias formas carnívoras e
herbívoras, uma das quais a dos cinodontes. Esses últimos desenvolveram altas
taxas metabólicas, o que possibilitava um estilo de vida ativo; aumento da
musculatura da mandíbula, o que possibilitava mordidas mais fortes; palato ósseo
secundário, que permitiu que esses animais respirassem enquanto se alimentavam;
e diferentes modificações no esqueleto, tornando-os mais ágeis. A terceira
linhagem de sinapsídeos, os mamíferos, teria se diversificado dos cinodontes e
os primeiros mamíferos provavelmente surgiram no final do período Triássico,
porém atingindo seu pico apenas na Era Cenozóica (~65 m.a.).
Os primeiros mamíferos do Triássico provavelmente
eram pequenos, do tamanho de ratos ou musaranhos, possuíam crânios avantajados,
maxilas cortantes e apenas duas dentições, uma decídua e outro permanente (difiodontia).
Esses mamíferos muito possivelmente também eram endotérmicos, embora sua
temperatura corpórea fosse mais baixa que a dos mamíferos atuais. Quanto aos pêlos,
eles provavelmente representam uma sinapomorfia de mamíferos, embora os pêlos geralmente
não são preservados no registro fóssil. Estudos relativamente recentes sugerem
que mamíferos possuem uma característica anatômica do ducto nasal-lacrimal, o
qual secretaria uma substância oleosa que é utilizada para vedar os pêlos (animal
espalha com as patas). Esta característica foi encontrada em um dos mais
antigos mamíferos conhecidos, os Morganucolom.
Contudo, ainda existe o argumento de que os cinodontes poderiam ter pêlos, uma
vez que tinham as taxas metabólicas mais altas e provavelmente necessitavam
desse anexo tegumentar para isolamento térmico.
Os mamíferos são geralmente classificados como
Allotheria (extintos), Prothoteria (monotremados) e Theria, a qual se divide em
Metatheria (marsupiais) e Eutheria (placentários). Os mamíferos atuais,
monotremados, marsupiais e placentários, se diversificaram no Terciário, e hoje
possuem cerca de 6500 espécies descritas. Estas espécies estão incluídas em 21
ordens, sendo uma dos monotremados (ovíparos), outra dos marsupiais (presença
dos marsúpios) e as demais dos eutérios (placentários). Elas ocorrem nos
ambientes aquáticos, terrestres e aéreos de todo o planeta. Possuem corpo com
nadadeiras (golfinho), membros anteriores modificados para o voo (morcego), variam
de 1,5 grama (morcego, Craseonycteris
thonglongyai) a 130 toneladas (baleia azul, Balaenoptera musculus), possuem uma ampla
variedade de dentição e sistemas digestórios, dependendo de seus modos
alimentares (herbívoros pastadores e podadores, roedores, ruminantes,
carnívoros e onívoros), alguns desenvolveram a ecolocação (cetáceos), alguns
têm glândulas mamarias sem mamilo e sem dentes na fase adulta (monotremados),
alguns com bolsas abdominais onde criam os filhotes (marsúpio), entre outras.
Portanto, os mamíferos são altamente diversificados em forma, tamanho/peso e
função.


Morfologia interna de um mamífero
Os mamíferos geralmente são animais de hábito
noturno, com algumas espécies solitárias que se encontram apenas nos períodos
reprodutivos, mas boa parte é social. Tanto é que eles são considerados os vertebrados
mais sociais. Há territorialidade, grupos com líderes (e.g. primatas) e o
cuidado parental é desenvolvido em praticamente todos eles, especialmente pelas
fêmeas que alimentam os filhotes com leite após o nascimento. As fêmeas também possuem
um ciclo estral periódico, quando se tornam receptivas e, geralmente após
rituais elaborados de corte, ocorre a cópula. A fecundação é sempre interna,
ocorrendo na maior parte dos casos a viviparidade, embora a oviparidade possa
ocorrer (e.g. ornitorrincos). A viviparidade pode ocorrer com as fêmeas dando a
luz a embriões, os quais são mantidos no marsúpio e têm amamentação estendida.
Pode ocorre tambem a viviparidade em que os filhotes ficam na placenta e nascem
completamente formados. A quantidade de filhotes varia, mas geralmente quanto
maior o animal menor a quantidade de filhotes. Alguns camundongos produzem 17
ninhadas de quatro a nove filhotes por ano. Enquanto, elefantes produzem um
filhote por geração, sendo quatro filhotes durante toda a vida que pode chegar
a 50 anos. Machos da mesma espécie podem matar e se alimentar dos filhotes a
fim de que as fêmeas se tornem receptivas mais rapidamente (e.g leões). Isso
geralmente ocorre quando um novo líder assume o grupo.
De maneira geral, os filhotes e os mais velhos são
os mais vulneráveis a predação, sendo os principais inimigos naturais dos
mamíferos, outros mamíferos, répteis, aves e em alguns casos até invertebrados (e.g
aranhas que se alimentam de pequenos camundongos). Por outro lado, muitos
mamíferos estão entre os principais predadores dos ecossistemas em que vivem,
sendo importantes reguladores das populações de outros animais.
Fonte: maior parte –
Pough et al.
A vida dos vertebrados. 4ª edição. São Paulo: Atheneu Editora, 2008.
Hickman et al. Principios integrados de zoologia. 11ª edição. Rio de
Janeiro: Guanabara Koogan, 2004.
Poucos detalhes –
Storer et al. Zoologia Geral. 6ª
edição. São Paulo: Companhia Editora, 2003.
Atividades - em breve!
Nenhum comentário:
Postar um comentário